The SpudCell and the Border of Life
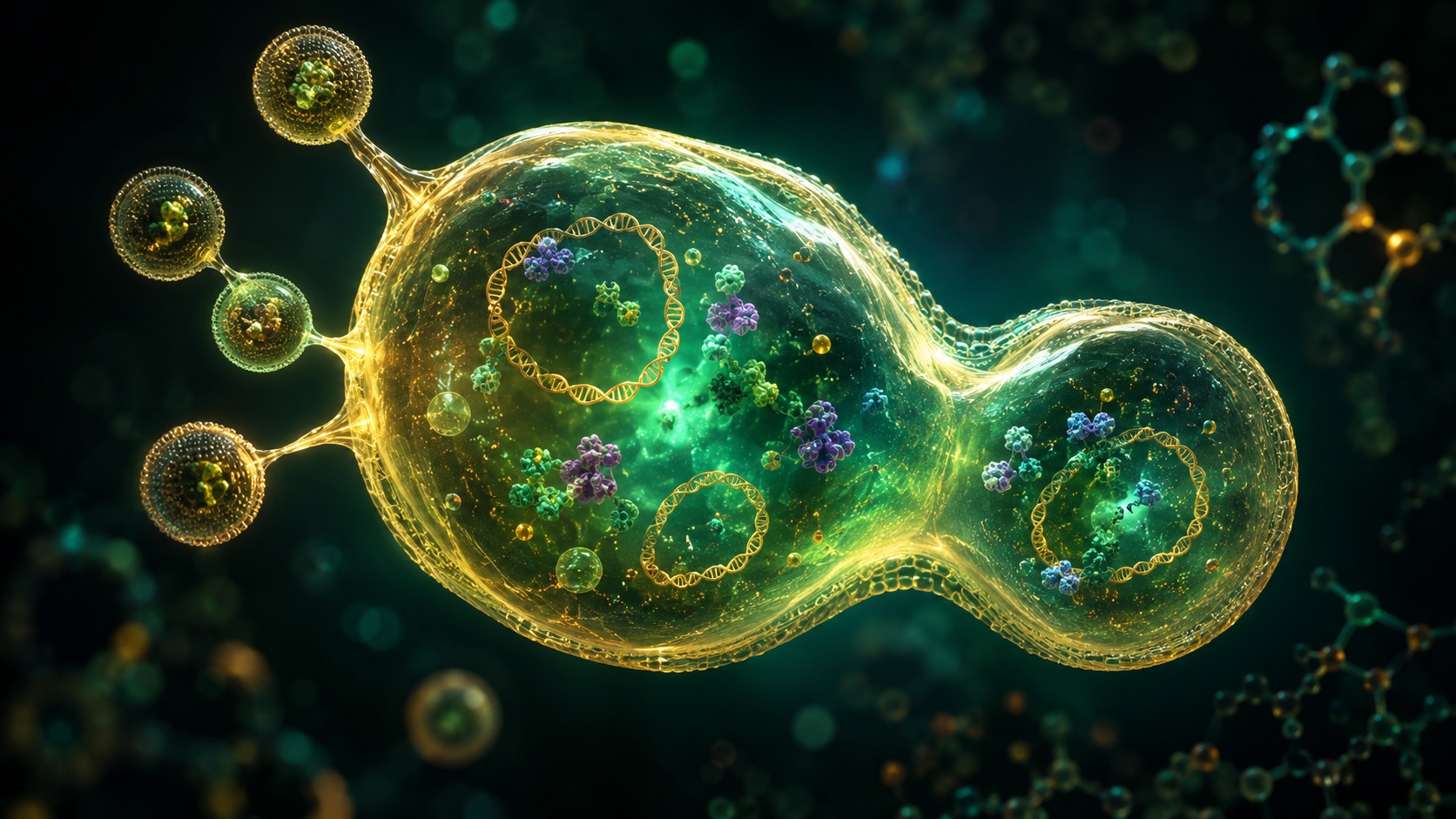
The SpudCell and the Border of Life: How a Potato-Shaped Droplet is Redrawing the Line Between Chemistry and Biology
The Shape of a Question
I did not expect to care about a potato.
Not the tuber itself, mind you, though I have strong opinions on roast potatoes and the tragic decline of the chip shop. No—I mean a microscopic, membrane-bound droplet that happens to look, under fluorescence microscopy, like a spud. A SpudCell. A thing assembled from purified, non-living biomolecular components that somehow manages to acquire materials, grow, copy its genetic material, and divide. It is not alive, or so the scientists insist, yet it performs several of the most fundamental dances of life with unsettling competence.
It is, in other words, a paradox wrapped in a lipid bilayer. And I find myself unable to stop thinking about it.
The question that haunts me—the question that should haunt anyone who reads the announcement from the University of Minnesota in July 2026—is deceptively simple: where exactly does chemistry end and life begin? We have grown accustomed to treating that boundary as a wall, a bright line, a metaphysical checkpoint with stern guards demanding to see your metabolic passport. But the SpudCell, that lumpy, self-replicating droplet, does not respect borders. It seeps across them. It makes a mockery of our categories. And in doing so, it forces us to confront something far larger than a single experiment: it forces us to ask what we mean when we say something is alive.
This is the story of how a team of researchers, led by Kate Adamala and Aaron Engelhart, built something unprecedented from the bottom up. It is also the story of why that something matters—not just to biologists in white coats, but to anyone who has ever wondered about the origins of life, the future of medicine, or the strange, stubborn persistence of biological organization in a universe that seems to prefer entropy.
The results were posted as a preprint in July 2026 and had not yet undergone formal peer review at the time of writing. That qualification matters. Extraordinary claims do not become less extraordinary merely because they arrive with attractive microscopy images and a university press release. But neither does the absence of peer review make the experiment uninteresting. It makes the story unfinished.
It is a story about lipids and proteins and DNA plasmids, yes, but it is also a story about ambition, limitation, and the hubris of thinking we can build what billions of years of evolution produced without consulting us.
Most of all, it is a story about a potato.
Let us begin.
The Long Road to Building a Blob
To understand why a potato-shaped droplet caused such a stir in the summer of 2026, one must first understand the history of synthetic biology, which is, in many respects, the history of human beings trying to play God with increasingly sophisticated chemistry sets. The field has always been split between two philosophical approaches, two temperaments, two distinct flavors of scientific audacity.
There is the top-down approach. This is the path of reduction, of stripping away. You take a living cell—something already alive, already complex, already humming with metabolic activity—and you pare it back, gene by gene, component by component, until you arrive at something approaching the minimal set of parts necessary for cellular life.
It is surgery. It is editing. It is the biological equivalent of taking a fully functional Swiss watch and removing gears until you find the smallest mechanism that still ticks.
The most famous exemplar of this approach came in 2010 from the J. Craig Venter Institute, where researchers constructed a synthetic version of the genome of Mycoplasma mycoides and transplanted it into a recipient Mycoplasma capricolum cell. The resulting organism, known as JCVI-syn1.0, was controlled by the chemically synthesized genome.
It was a triumph. It was also, in a sense, cheating.
The membrane, ribosomes, cytoplasm, and metabolic infrastructure of the recipient cell were already present. The genome was synthetic, but the initial chassis was borrowed. It was rather like replacing every page in a railway timetable and announcing that one had built a railway.
Then there is the bottom-up approach. This is the path of creation ex nihilo, or as close to nothing as chemistry permits. You start not with a living cell but with purified components—lipids, nucleic acids, proteins, salts, sugars, enzymes, and molecular machinery—and attempt to assemble them into something that behaves like life.
No pre-existing intact cell. No inherited cellular chassis. Just molecules, a flask, and a prayer.
This is alchemy. This is the origin-of-life research program dressed in modern lab coats. And for decades, it produced fascinating curiosities—protocells that could encapsulate RNA, vesicles that could divide under certain conditions, and systems that could reproduce particular cellular functions—but nothing that combined growth, genetic replication, division, and competition within a single engineered platform.
Nothing that felt, even from a distance, quite so cell-like.
Until the SpudCell.
The University of Minnesota announced the work on July 1, 2026, and the corresponding preprint appeared shortly afterward. Adamala and Engelhart, working with colleagues at the University of Minnesota Twin Cities, had constructed a synthetic cellular system from purified biomolecular components that could perform an unusually broad collection of life-like behaviors: it could receive packaged molecular supplies, grow, copy its genetic instructions, and divide into daughter cells.
It could also undergo competition and selection. Researchers constructed variants with different growth characteristics, placed them together, and watched the faster-growing form become more abundant over five generations.
The New York Times described the system as possessing many of the hallmarks of life. John Glass of the J. Craig Venter Institute, no stranger to minimal cells himself, called the work a landmark event in the history of biology.
And yet.
And yet the creators themselves stopped short of calling it alive.
This tension—between what the SpudCell does and what it is—will occupy us for the remainder of this journey. But first, we must look inside the droplet.
What Lies Within: Anatomy of an Almost-Life
Imagine, if you will, a microscopic bubble. Not a soap bubble—that would be too fragile, too ephemeral—but a bubble made of lipids, those greasy, water-fearing molecules that naturally arrange themselves into membranes when placed in aqueous environments.
This is the lipid bilayer, the familiar architecture of most cellular boundaries, and it can form spontaneously, without genetic guidance, as if chemistry has a prefabricated fondness for compartments.
The SpudCell begins here: with lipids enclosing a droplet of chemical broth.
That broth is not simple. It contains roughly one hundred types of purified proteins and small molecules, carefully selected and mixed like the ingredients of an impossibly complex consommé. Among these are ribosomes, the molecular machines that read genetic instructions and assemble proteins.
These ribosomes are not synthesized by the SpudCell. They are supplied by the researchers, having ultimately been derived from existing biology, and this is the first of several dependencies that prevent the system from being fully autonomous.
The SpudCell cannot build its own protein factories. It inherits them, like a trust-fund cell living off an endowment it did not earn.
The system also uses a cell-free protein-production mixture known as the PURE system. This includes approximately thirty-six purified enzymes, along with ribosomes, transfer RNAs, amino acids, and other components needed to translate genetic instructions into proteins.
This number should not be confused with the number of genes in the SpudCell genome. There happen to be thirty-six of those too, because nature and laboratory engineering occasionally enjoy the small joke of reusing a number.
The genome is not a single, elegant chromosome like the chromosomes found in natural cells, but seven separate DNA plasmids—small, circular pieces of genetic material—totaling approximately ninety thousand base pairs and encoding thirty-six genes.
To put this in perspective, the human genome contains roughly three billion base pairs. Even the smallest independently replicating bacterial cells possess genomes several times larger than that of SpudCell. The engineered minimal bacterium JCVI-syn3.0, for example, carries approximately 531,000 base pairs and 473 genes.
Some theoretical estimates have suggested that an exceptionally stripped-down genetic system might require around 113,000 base pairs. SpudCell appears to operate below even that provocative figure, but the comparison is imperfect. A natural cell must encode and manufacture much of its own machinery. SpudCell survives only because the surrounding laboratory supplies ribosomes, enzymes, energy sources, transfer RNAs, membrane materials, and other molecular necessities.
Its genome is small partly because the unpaid labor has been outsourced.
The genome is fragmented, modular, almost provisional. It is less a blueprint than a set of sticky notes with instructions scrawled in haste.
But those instructions are sufficient, barely, to orchestrate a remarkable sequence of events. The SpudCell receives nutrients and ready-made molecular machinery from specially designed feeder liposomes. Its supplied ribosomes translate genetic code into functional proteins. It grows, swelling like a satisfied amoeba.
And then, most remarkably, it divides.
Here is where the SpudCell diverges from many natural cells in a way that is both elegant and slightly unsettling. Natural cellular division often depends on cytoskeletal machinery—organized protein structures that constrict, reshape, or physically separate the cell.
Building a reliable cytoskeleton from scratch has been one of the great bottlenecks in synthetic-cell research. How does one engineer a self-organizing structural network without billions of years of evolutionary tinkering, breakage, compromise, and repair?
The SpudCell sidesteps much of this problem. Instead of constructing an elaborate internal division machine, it expresses membrane-crowding proteins that accumulate at the lipid surface. Their presence alters the physical properties of the membrane and helps generate the deformation necessary for division.
The process is not entirely self-sufficient. Externally supplied streptavidin and linker molecules assist the membrane rearrangement. The division is therefore genetically directed but environmentally assisted.
It is division by congestion, with help from the laboratory staff.
The cell does not simply pinch itself in half. It changes the conditions of its own membrane until splitting becomes physically favorable.
This is engineering as improvisation. It is biology by way of jazz.
And it works. The SpudCell completes a laboratory cycle of growth, genome replication, and division. Under selective conditions, variants expressing greater quantities of particular proteins can obtain more feeder material, grow faster, and produce more descendants.
After five generations, the faster variant dominates the population.
The system therefore demonstrates heredity and selection among deliberately engineered variants, two essential ingredients of Darwinian evolution. But the useful variation was designed by the researchers rather than generated through spontaneous mutation.
The experiment is thus not yet a complete demonstration of autonomous evolution. It is closer to a rehearsal in which the actors can perform the scene, but the playwright remains in the room.
And the performance lasts only five to ten generations.
Then the borrowed ribosomes and other inherited molecular components degrade or become diluted. The lineage sputters and dies. The genome, fragmented across seven plasmids, is not reliably transmitted intact to every offspring. After several generations, many daughter cells no longer possess the complete genetic set.
The system collapses under the weight of its own dependencies.
It is, in the end, a brilliant failure. Or a failed brilliance. The distinction depends on how generous you are feeling.
The People Who Built the Border
Science is never a disembodied enterprise, though we often write about it as if discoveries emerge from the ether fully formed. They do not. They emerge from particular minds in particular places at particular moments, and the SpudCell is no exception.
Kate Adamala, an associate professor at the University of Minnesota’s College of Biological Sciences, has spent her career probing the boundary between chemistry and biology. Her work sits at the intersection of synthetic biology and origin-of-life research, two fields that ask essentially the same question—how does non-living matter become living?—from opposite directions.
Adamala approaches this question with the temperament of an engineer and the curiosity of a philosopher. She does not merely want to understand life; she wants to build simplified versions of it, piece by piece, until the understanding emerges from the construction.
Aaron Engelhart, her collaborator and a faculty member at the University of Minnesota, brings complementary expertise in molecular engineering, nucleic acids, and synthetic biology.
The study’s first author was Nathaniel J. Gaut, working with Christopher Deich, Brock Cash, Tanner Hoog, Engelhart, and Adamala. The ordering of names in a scientific paper is its own peculiar social machinery, one that can make a team effort look like a queue at a railway counter. But it matters, and credit should follow the paper rather than the press release.
Together, the researchers spent approximately five researcher-years—a unit of measurement that should be standardized, like light-years but for academic suffering—developing the system.
The work was grueling. The knowledge was difficult to transfer; collaborators sometimes required in-person demonstrations to reproduce particular techniques. This is not yet how scalable engineering works. This is artisanal cell-building, closer to bespoke tailoring than industrial manufacturing.
The rejection of an early version of the paper by the journal Cell—reportedly after one reviewer described the system as “not real biology”—adds a layer of irony, though perhaps not quite the morality play it first appears to be.
Here was a system that could grow, replicate DNA, and divide, dismissed in part because it did not fit comfortably within a conventional category of biology. But extraordinary claims about synthetic life also deserve unusually careful scrutiny, especially when the work has not yet been independently reproduced or formally peer reviewed.
The reviewer may have been defending a category too rigidly. Or the reviewer may have been asking whether a heavily supported molecular construct should be judged by the same standards as an autonomous cell. Possibly both things were true at once, which is how science generally arranges its arguments when nobody is looking.
The episode captures a genuine disciplinary tension. Should a chemically assembled system be judged as cell biology, engineering, or both? Is a cell defined by what it does, by where its components came from, or by how thoroughly it can sustain itself without a laboratory hovering nearby with a pipette?
Adamala’s response to this challenge has been characteristically ambitious. Rather than retreating into proprietary secrecy, she and her collaborators launched Biotic, a public-benefit research and engineering institution dedicated to building shared technical infrastructure for synthetic-cell engineering.
The goal is to transform SpudCell from a bespoke curiosity into a chassis—a standardized, modular platform upon which other researchers can build.
This is the philosophy of open-source biology, and it represents a bet that the future of artificial life will be collaborative or it will not be at all.
The Machinery of Almost-Life: How It Works
To truly appreciate the SpudCell, one must understand the molecular machinery that makes its behaviors possible. This requires a brief descent into the biochemical weeds, though I promise to provide a ladder.
Let us begin with the central dogma of molecular biology, a phrase that sounds like religious doctrine and functions with similar authority.
In cellular life, genetic information usually flows from DNA through RNA to protein, although biology has accumulated several important variations on this route. DNA is the archive, the library of instructions. Messenger RNA is the photocopied page carried from the stacks to the workshop. Protein is often the product, the tool or machine that performs much of the work of the cell.
This flow is not a complete description of biology. Some RNAs perform functions without becoming proteins. Retroviruses copy RNA back into DNA. Gene regulation twists the simple arrow into something resembling the Calcutta tram map after a thunderstorm.
But the DNA-to-RNA-to-protein pathway remains one of the central organizing principles of cellular life.
The SpudCell operates on it too, which is one reason it feels so uncannily alive.
The production of proteins requires translation, the process by which RNA messages are decoded into amino-acid sequences. This is the job of the ribosome, a molecular machine of staggering complexity, composed of both protein and RNA, that reads the genetic code and stitches amino acids together in the correct order.
Natural cells build their own ribosomes through an elaborate, multi-step process involving many genes, RNAs, enzymes, and assembly factors.
The SpudCell cannot do this.
It is, in this respect, like a brilliant novelist who cannot manufacture a pen. It can write, but only if someone provides the instrument.
The supplied ribosomes are functional but finite. They degrade. They are diluted between daughter cells. They are not replaced by newly manufactured ribosomes.
This is one reason the lineage dies after five to ten generations: the protein factories wear out, and there are no new factories to take their place. It is a fatal dependency, like a civilization that knows how to use computers but not how to build them, collapsing when the last server overheats.
Then there is metabolism, the network of chemical reactions through which living cells obtain energy, transform raw materials, maintain their internal environment, and manufacture cellular components.
Here the SpudCell is far less autonomous than an ordinary living cell.
It does not possess a complete, self-sustaining metabolic network. Instead, the researchers feed it specially prepared liposomes containing sugars, lipids, ribosomes, enzymes, transfer RNAs, and other ready-made molecular components. The SpudCell fuses with these feeder packages and acquires their contents.
It does not so much grow its own wheat as receive parcels from a highly attentive grocery service.
This shortcut is one reason the genome can remain so small. Many of the functions that a natural cell would have to encode, regulate, repair, and reproduce are performed outside the SpudCell and delivered to it in finished form.
It can process some inputs and use them to produce proteins, copy DNA, expand its membrane, and divide. But it cannot independently construct the complete molecular economy upon which those activities depend.
The genome replication is another marvel of minimalism. The seven plasmids must be copied before division, and this copying is carried out using a replication system based on supplied enzymes, including a DNA polymerase derived from bacteriophage Phi29.
But because the genome is fragmented, inheritance is messy. Daughter cells do not reliably receive complete copies of all seven plasmids. Genetic information is lost, diluted, or unevenly distributed.
It is heredity without fidelity, reproduction without dependable continuity.
The SpudCell replicates, but it does not replicate well.
And yet.
And yet the system does replicate its DNA. It does grow. It does divide. It does permit experimentally constructed variants to compete.
Under conditions in which feeder materials are limited, a variant that produces more of the relevant membrane protein can acquire more supplies, grow more rapidly, and become increasingly common.
The SpudCell therefore demonstrates that several core behaviors associated with life—compartmentalization, protein synthesis, genome replication, growth, division, heredity, and selection—can be coordinated within a chemically assembled system, even if that system remains fragile, externally supported, and temporary.
It is not a living cell.
But it is a life-like cell.
And the difference between those two categories, once assumed to be a chasm, now looks suspiciously like a gradient.
Where This Happens: From the Lab Bench to the Imagination
The SpudCell was born in laboratories at the University of Minnesota, in buildings no doubt filled with the hum of freezers, the gleam of pipettes, and the faint, persistent smell of disinfectant.
But its significance extends far beyond that specific place.
It belongs to a broader landscape of synthetic-biology laboratories scattered across the globe, from the J. Craig Venter Institute in California to research groups in Europe, Japan, and elsewhere. It is part of an emerging ecosystem of artificial-life research that is increasingly collaborative, interdisciplinary, and open.
The physical location matters less than the conceptual space the SpudCell occupies.
It sits at the intersection of multiple disciplines: synthetic biology, which seeks to engineer biological systems; origin-of-life research, which seeks to understand how life emerged from non-living chemistry; systems biology, which seeks to understand how complex behaviors emerge from interacting components; and bioengineering, which seeks to harness biological processes for practical applications.
It is a node in a network of ideas, a point where multiple intellectual trajectories converge.
Its possible applications, should the technology mature over many years or decades, could be substantial.
Imagine custom cells designed to produce therapeutic molecules that natural biology cannot easily synthesize, perhaps incorporating amino acids that evolution never adopted. Imagine bioremediation systems designed to capture particular pollutants or manufacture useful chemicals under mild conditions. Imagine artificial compartments that act as programmable diagnostic devices, sensing molecular signals and responding in controlled ways.
The SpudCell is not any of these things yet.
It does not manufacture medicines in hospitals, patrol arteries, clean rivers, grow organs, or solve climate change before lunch. It is a fragile research system that requires expert preparation and survives for a limited number of generations.
The path from such a laboratory construct to useful technology is long, uncertain, and cluttered with the skeletons of previous revolutions.
But SpudCell is a first draft of a possible platform.
This is why the open philosophy of Biotic matters. If synthetic cells are to become an engineering discipline rather than a laboratory curiosity, they will require standardization, modularity, reproducibility, and shared infrastructure.
Adamala has argued that engineering disciplines depend on modular systems and that foundational modules should be developed openly rather than controlled as private toll roads.
The SpudCell is intended as a chassis.
Whether anyone can turn it into a reliable vehicle remains to be seen.
Why Any of This Matters: The Philosophy of the Potato
I promised you would care about this potato-shaped droplet, and now I must deliver on that promise.
The SpudCell matters on multiple levels, some practical and some profound.
On the practical level, it represents a proof of concept. It demonstrates that several fundamental behaviors associated with life—growth, genetic replication, division, heredity, and selection—can be coordinated in a system assembled from purified components rather than inherited as part of an already living cell.
This is not trivial.
For decades, the bottom-up approach to synthetic biology was plagued by the suspicion that life might require something irreducible, some vital spark, some élan vital that could not be manufactured.
The SpudCell does not fully eliminate that suspicion because it still depends heavily on components produced by living organisms. Its ribosomes, enzymes, transfer RNAs, and other molecular machinery are not conjured from elementary chemicals.
Nevertheless, it suggests that life-like organization can arise from mechanism rather than magic.
There is no visible vital force hiding in the membrane. There are molecules, physical constraints, information, energy, and a staggering amount of careful laboratory work.
This has implications for origin-of-life research, though they must be stated carefully.
The SpudCell is not a plausible model of Earth’s earliest cell. It is far too elaborate, far too dependent on modern proteins, purified enzymes, ribosomes, synthetic DNA, and sophisticated laboratory intervention.
No prehistoric pond contained a refrigerated PURE protein-expression kit, unless the history of the planet is considerably stranger than geologists have admitted.
But the system is still useful as a proof of principle. It shows that compartmentalization, genetic information, molecular synthesis, growth, and division can be coupled within a designed chemical environment.
By constructing such systems and observing where they fail, researchers may learn which cellular features are indispensable, which can be replaced, and which are merely solutions evolution happened to discover first.
On the philosophical level, the SpudCell challenges our definitions.
What is life?
The question is ancient, and the answers have always been unsatisfying.
Movement? Crystals and flames can spread.
Metabolism? Fire consumes fuel and releases energy.
Reproduction? Computer viruses copy themselves.
Heredity? Prions transmit structural information.
Evolution? Populations of molecules can sometimes undergo selection without possessing anything resembling a cell.
The SpudCell forces us to confront the inadequacy of simple checklists. It is too organized to be dismissed as ordinary chemistry, yet too dependent to be accepted without qualification as biology.
It occupies a liminal space, a borderland, and its existence makes that borderland harder to ignore.
There is also a subtler, more psychological reason the SpudCell matters.
It is a mirror.
In building something that approximates life, we are forced to examine what we value about life itself.
Is it autonomy? The SpudCell lacks much of that.
Is it continuity? Its lineages are fleeting.
Is it complexity? It is deliberately simplified.
Is it the capacity to evolve? It permits heredity and selection, but not yet open-ended evolution driven by spontaneously arising variation.
Is it the ability to maintain and recreate the machinery required for one’s own existence? By that measure, SpudCell remains dependent on a world outside itself.
The SpudCell strips away some of the grandeur of biology and reveals the underlying mechanics. In doing so, it tempts us to ask whether we are more than mechanics ourselves.
This is not comfortable.
It was never meant to be.
The Misconceptions We Carry
No discussion of artificial life would be complete without addressing the misconceptions that cluster around it like barnacles on a hull.
The first and most persistent is the belief that the SpudCell is unquestionably alive.
It is not, or at least not by the standards most biologists would apply to an autonomous organism.
It cannot maintain itself indefinitely. It cannot synthesize its own ribosomes. It cannot manufacture all the enzymes and transfer RNAs upon which it depends. Its genome is too fragmented to ensure faithful inheritance across many generations. It requires carefully prepared feeder liposomes and externally supplied molecular assistance.
It is less a fully independent organism than a chemically assembled performance of selected cellular functions.
But this raises a second misconception: that the distinction between “alive” and “not alive” must therefore be sharp and immutable.
The SpudCell suggests otherwise.
It suggests that the boundary may be porous, gradational, and dependent on which properties we consider essential. We classify things as living or non-living because the distinction is useful. The universe is under no contractual obligation to arrange its chemistry according to our dictionaries.
A third misconception is that the SpudCell represents an immediate threat or promise.
It does not.
This is early-stage science, fragile and finicky, requiring years of expert work to produce a system that persists for only a handful of generations.
The path from laboratory curiosity to practical application is long, uncertain, and littered with the remains of overhyped technologies. The SpudCell is not about to revolutionize medicine tomorrow.
It might not revolutionize medicine at all.
But it opens a door, and what lies beyond that door is worth investigating.
A fourth misconception is that the system has demonstrated full Darwinian evolution.
It has not.
Researchers constructed variants with different levels of a growth-related protein and demonstrated that one could outcompete another. This is genuine selection acting on heritable, engineered differences.
But the advantageous variation did not arise spontaneously through mutation, and the system has not demonstrated indefinite, open-ended evolution.
It possesses some of evolution’s machinery, but the laboratory still holds the steering wheel.
Finally, there is the misconception that this work is solely about utility.
It is not.
It is also about understanding.
The SpudCell is a tool for probing the minimal requirements of cellular behavior. By building simplified systems and observing where they fail, scientists can identify which components are truly essential, which can be replaced by engineered shortcuts, and which may be historical baggage accumulated over billions of years.
It is reductionism as revelation, engineering as epistemology.
The Technologies That Made the Potato Possible
Behind the SpudCell lies a constellation of technologies, methods, and discoveries that made its creation possible.
It did not emerge from a vacuum. It stands on the shoulders of giants, some of whom did not live to see their work bear this particular fruit.
Lipid self-assembly, the spontaneous formation of membrane-bound vesicles from fatty molecules, was characterized in increasing detail during the twentieth century. This phenomenon, which can occur without genetic instruction or metabolic energy, provided the foundational insight that cellular compartments could form from relatively simple chemistry.
The SpudCell’s membrane is a direct descendant of this understanding.
Cell-free protein synthesis, the ability to produce functional proteins from genetic templates without intact living cells, matured during the late twentieth and early twenty-first centuries.
The SpudCell uses a purified cell-free expression system containing ribosomes, enzymes, transfer RNAs, amino acids, energy sources, and other molecular components. This allows it to convert genetic information into functional proteins while avoiding many of the complexities of an intact natural cell.
Synthetic genomics, the design and construction of artificial genetic sequences, was pioneered by researchers including Craig Venter, Hamilton Smith, Daniel Gibson, and many others.
Their work on synthetic bacterial chromosomes and minimal genomes demonstrated that DNA could be designed computationally, chemically synthesized, assembled, and used to control cellular behavior.
The SpudCell’s seven-plasmid genome, though tiny by biological standards, is a product of this capability.
Isothermal DNA amplification and replication systems, including enzymes derived from bacteriophage Phi29, made it possible to copy DNA within the synthetic compartment without reproducing the entire replication machinery of a natural bacterium.
Membrane engineering allowed the researchers to control how SpudCells received supplies, expressed pore-forming and membrane-associated proteins, expanded their membranes, and divided.
DNA sequencing and synthesis technologies, whose costs have fallen dramatically over the past several decades, made it feasible to design, build, test, and revise genetic systems with a speed that would once have been financially absurd.
The SpudCell would have been prohibitively difficult to attempt in 2006.
In 2026, it was merely extraordinarily difficult.
Modern synthetic-cell research more broadly also benefits from computational modelling, microscopy, microfluidics, automated liquid handling, and bioinformatics. Not every one of these technologies was necessarily central to the reported SpudCell experiment, but together they form the technical landscape in which such work has become conceivable.
The SpudCell is therefore not merely a product of biology.
It is a product of chemistry, physics, engineering, computation, manufacturing, and the human tendency to make small containers and then ask what might happen inside them.
The Limitations That Define It
For all its brilliance, the SpudCell is defined as much by what it cannot do as by what it can.
These limitations are not incidental.
They are instructive.
They point toward the frontiers that remain.
The ribosome dependency is the most glaring. Ribosomes are among the most complex molecular machines in nature, and the SpudCell cannot build them.
This is not a minor inconvenience. It is a fundamental incapacity.
A cell that cannot manufacture its own protein-synthesis machinery is like a society that cannot educate its own engineers. It can function for a time, but it cannot indefinitely recreate the infrastructure upon which its survival depends.
Solving this problem—building a synthetic cell that can produce and assemble its own ribosomes from genetic instructions—would be a leap comparable to the SpudCell itself, perhaps greater.
The fragmented genome is another critical weakness.
Natural cells typically carry their genetic information on one or a few chromosomes that are copied and distributed through organized segregation systems. The SpudCell’s seven plasmids are not reliably inherited as a complete set.
Daughter cells may receive incomplete genetic complements, leading to functional decline over successive generations. Reports from the project indicate that after five generations, only a minority of descendants still retain the complete plasmid set.
Consolidating the genome into a more stable chromosome, or engineering a reliable segregation mechanism, is therefore a priority. It is also a non-trivial engineering challenge.
The lack of autonomous metabolism restricts the SpudCell to an environment constructed specifically for it.
Natural cells have evolved elaborate metabolic networks that allow them to extract energy, synthesize components, maintain chemical balances, repair damage, and survive environmental change.
The SpudCell depends on packaged molecular supplies prepared by researchers. It is a specialist in a world that has been custom-built around its limitations.
The externally assisted division process is another dependency. Although the system genetically produces membrane-associated proteins involved in reshaping the vesicle, its division still relies on externally supplied linking molecules and carefully controlled conditions.
And then there is the temporal limitation.
Five to ten generations is not a lineage.
It is a flicker.
It is a proof of concept that burns out before it can become a proof of persistence.
For the SpudCell to become a durable platform for synthetic biology, it must close more of the loops of self-maintenance. It must manufacture or renew its own machinery, preserve its complete genome, regulate its internal environment, and continue reproducing without the molecular equivalent of room service.
These limitations are not failures.
They are the agenda.
They are the map of the next decade’s research.
Stepping Back: The View from the Border
I find myself returning, again and again, to the image of that potato-shaped droplet, suspended in its chemical broth, dividing through membrane deformation, competing for feeder materials, and dying when its borrowed machinery fails.
It is a humble thing.
It is also a revolutionary thing.
And the tension between those two truths is, I think, the essence of what the SpudCell represents.
We are living through a moment when the boundary between the natural and the artificial is dissolving—not with a dramatic rupture, but with a slow, steady seepage.
We have grown accustomed to artificial intelligence that thinks, after a fashion. Now we are confronted with artificial cells that live, after a fashion.
The questions they raise are not new, but they are newly urgent.
What do we owe to something that behaves like life but was built by human hands?
What does it mean to create when the thing created approximates the fundamental processes of biology?
Are we playing God, or merely discovering that God’s workshop contains more physical chemistry and fewer thunderbolts than expected?
I do not know the answers.
I suspect the answers are less important than the asking.
The SpudCell is not alive in the ordinary sense.
But it is alive-adjacent.
It is life-like.
It is a ghost in the machine, except the machine is made of lipids and the ghost is made of chemistry.
And in that liminal space, that borderland between the living and the non-living, we find some of the most profound questions science can ask. Not because they have simple answers, but because the asking reshapes the categories with which we understand ourselves.
Kate Adamala and her colleagues have given researchers a possible chassis.
The rest—scientists, engineers, philosophers, regulators, and citizens—must decide what can responsibly be built upon it.
The SpudCell can grow, copy its DNA, divide, and compete for several generations before its inherited machinery decays and its fragmented genome falls apart.
Perhaps its descendants, biological or intellectual, will persist much longer.
Perhaps they will eventually become autonomous.
Perhaps they will remain elegant laboratory contraptions, useful chiefly because their failures reveal what natural cells quietly accomplish every second.
That, too, would be fitting.
We have always been a species that builds things and then wonders what they mean.
The SpudCell is only the latest, and perhaps one of the most humbling, entries in that long catalog of human creation.
It is a potato.
It is a mirror.
It is a question that divides, and in dividing, multiplies.
And I, for one, cannot look away.
P.S. References and Further Reading
Gaut, Nathaniel J., Christopher Deich, Brock Cash, Tanner Hoog, Aaron E. Engelhart, and Katarzyna P. Adamala. “A Chemically Defined Synthetic Cell Capable of Growth and Replication.” bioRxiv, posted July 2, 2026. https://doi.org/10.64898/2026.07.01.735724. Preprint; not yet peer reviewed.
Kupferschmidt, Kai. “Lab-Created ‘SpudCell’ Marks ‘Stunning’ Step Toward Building Life from Scratch.” Science, July 1, 2026.
Saplakoglu, Yasemin. “For the First Time, a Cell Built From Scratch Grows and Divides.” Quanta Magazine, July 1, 2026.
Zimmer, Carl, and Marco Hernandez. “This Cell Feeds, Grows and Reproduces. And It’s Manmade.” The New York Times, July 1, 2026.
University of Minnesota Twin Cities. “World’s First Synthetic Cell with a Complete Life Cycle Could Revolutionize Biological Engineering.” Research brief, July 1, 2026.
Biotic. “SpudCell: The First Synthetic Cell with a Complete Cell Cycle.” Project documentation, accessed July 11, 2026.
Gibson, Daniel G., et al. “Creation of a Bacterial Cell Controlled by a Chemically Synthesized Genome.” Science 329, no. 5987 (2010): 52–56. https://doi.org/10.1126/science.1190719.
Hutchison, Clyde A. III, et al. “Design and Synthesis of a Minimal Bacterial Genome.” Science 351, no. 6280 (2016): aad6253. https://doi.org/10.1126/science.aad6253.
Ricardo, Alonso, and Jack W. Szostak. “The Origin of Life on Earth.” Scientific American 301, no. 3 (2009): 54–61.
Word Cloud
